
И. А. ЕФРЕМОВ
О СТРОЕНИИ КОЛЕННОГО СУСТАВА ВЫСШИХ ДИЦИНОДОНТОВ
(Представлено академиком Е. Н. Павловским 23 I 1951)
Скелет аномодонта, найденный в 1936 г. П. И. Климовым в триасовых отложениях р. Донгуз в Чкаловской обл. и хранящийся в Палеонтологическом музее Академии наук СССР (№ 159/1 ПИН), был предварительно описан мною ( 1,2) как новый вид листрозавра — Lystrosaurus klimovi.
Новый аномодонт отличался своей сравнительно высокой организацией, позволившей с определенностью говорить о триасовом возрасте донгузских отложений. Наиболее характерным оказалось строение крестца из 8 позвонков, развитие большой грудины и мощного olecranon на локтевой кости.
В процессе дальнейшей препаровки выяснилось, что донгузская форма более близка к типичным дицинодонтам, должна быть исключена из рода Lystrosaurus и представляет собою новый род.
Систематическая характеристика донгузского дицинодонта следующая: отряд Anomodontia, сем. Dicynodontidae, род Rhadiodromus nov., вид klimovi Efr.
Диагноз должен быть дополнен описанием задней конечности, по характеру которой дано название нового рода (ra iwz — легко, без труда. dromaz — бегающий).
Правая задняя конечность Rhadiodromus сохранилась неполностью — отсутствуют верхняя часть бедра и большинство элементов стопы. Уцелевшая часть конечности отличается необычным для дицинодонтов строением коленного сустава (см. рис. 1, А и Б, рис. 2).
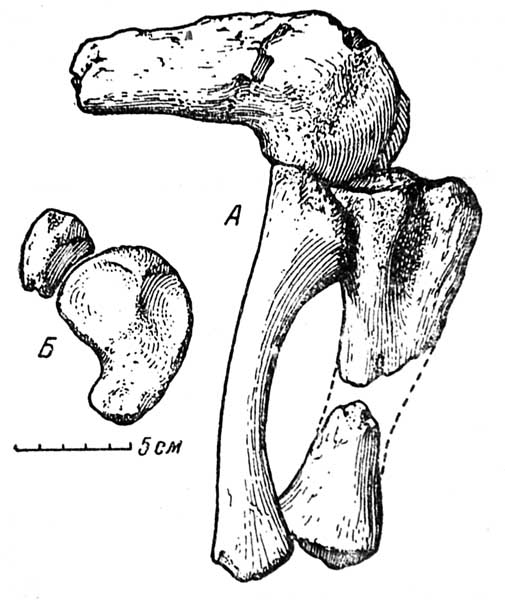
Рис. 1. Rhadiodromus klimovi gen. nov. A — бедро и голень в естественном положении, с наружной стороны; коленный сустав сильно согнут для показа размаха движения; Б — большая и малая берцовые кости сверху (суставные поверхности)
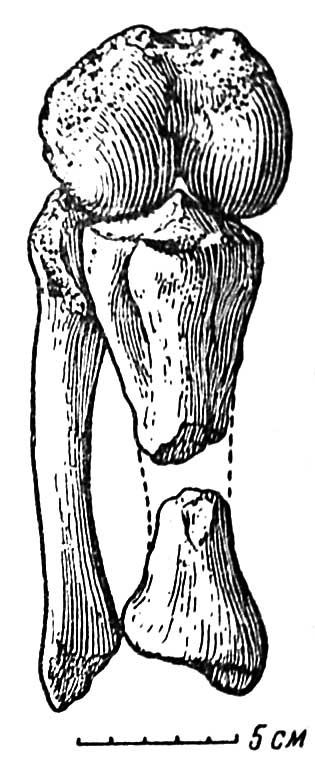
Рис. 2. Rhadiodromus klimovi gen. nov. Бедро и голень в естественном положении, спереди; коленный сустав сильно согнут для показа суставных поверхностей
Сравнительно длинная бедренная кость, выпуклая на спинковой поверхности и вогнутая на плантарной, обладает цилиндрическим, а не плоским, свойственным аномодонтам, сечением. Дистальный конец кости несет столь хорошо сформированную суставную поверхность, что кажется принадлежащим млекопитающему с окостеневшими эпифизами, а не древнему тероморфу.
Наружный и внутренний мыщелки развиты одинаково, разделены глубокой межмыщелковой вырезкой с fossa intercondyloidea в центральной части.
На наружной поверхности латерального мыщелка (см. рис. 1, А) отчетливо выражена связочная ямка и ямка подколенного мускула (fossa m. poplitei). Выше ямки развит пологий и широкий связочный бугор. Сходные образования более слабо выражены на медиальном мыщелке.
Гладкая блоковидная поверхность каждого мыщелка очерчивает правильную дугу больше полуокружности (около 200°), что доказывает большой размах движения блоковидного сустава бедренной кости.
Большая берцовая кость (tibia) выделяется своей сильно вытянутой в передне-заднем направлении верхней суставной площадкой. Подобная длинная суставная поверхность (см. рис. 1, А) характерна для подолгу передвигающихся копытных млекопитающих, например для овец.
Подобно бедру, суставные поверхности tibia, как верхняя, так и нижняя, очень хорошо окостенели. Межмыщелковая возвышенность проксимального конца хорошо развита (см. рис. 2). Чрезвычайно мощно развита tuberositas tibiae, переходящая ниже в очень сильно выступающий гребень (crista tibiae). Это говорит о мощном развитии передних связок коленного сустава, не меньшем, чем у млекопитающих, обладающих коленной чашкой. На латеральной стороне большой берцовой кости проходит очень глубокий мускульный жолоб, свидетельствующий о могучих разгибателях пальцев и большеберцовых мускулах, управляющих стопой и нужных для быстрого передвижения. На дистальном конце кости развита винтообразная суставная поверхность, далеко не столь сложная, как у млекопитающих, но хорошо сформированная.
Малая берцовая кость (fibula) — уплощенная, с менее хорошо окостеневшими концами. Она слегка искривлена в латеральном направлении, вследствие чего межкостное пространство (spatium interosseum) большое. Верхний конец fibula несет валикообразную суставную поверхность, изогнутую по широкой дуге, соответственно диапазону движения бедренного мыщелка (см. рис. 1, А).
Описанное строение коленного сустава Rhadiodromus резко отличается от более древних дицинодонтов из северодвинских раскопок Амалицкого, обладающих уплощенными и укороченными костями задней конечности. Бедро с слабо развитой головкой и дистальными мыщелками все же характерно своей относительно вертикальной ориентировкой и большим поворотом колена вперед, чем у других современных им рептилий, исключая цинодонтов. У громадных поздних дицинодонтов Южной Америки коленный сустав также направлен вперед, но бедро и тибия плоские, со скошенными суставами, явно утяжеленные и ограниченные в движении вследствие большого веса животного.
Rhadiodromus достигает «средних» размеров, примерно соответствующих величине крупного барана или небольшого бычка, и может обладать значительной подвижностью. Донгузскую форму следует рассматривать как наиболее подвижную из всех дицинодонтов. Высоко прогрессивное строение коленного сустава комбинируется с сравнительно короткой голенью, с большими размерами малой берцовой кости и, очевидно, плоской и широкой стопой. На этой примитивной основе развился коленный сустав, не менее совершенный, чем у многих примитивных млекопитающих. Такое приспособление в общем идет по пути приспособительной эволюции млекопитающих и противоположно архозаврам, у которых большой размах задней конечности осуществляется развитием интертарзального сочленения и аналога птичьей цевки.
Коленный сустав Rhadiodromus klimovi, допускающий значительный размах движения голени и направленный вперед, коррелятивно связан с развитием удлиненного таза на мощном крестце из 8 позвонков. К приспособлениям, обеспечивающим возможность интенсивного передвижения, можно отнести и olecranon локтевой кости. Однако передняя конечность Rhadiodromus более массивна и как бы противоречит подвижному строению задней. В такой противоречивой анатомической структуре предстает перед нами донгузский дицинодонт, биологически, несомненно, представлявший животное, передвигавшееся быстрее и продолжительнее своих более древних и крупных сородичей.
Подобное первоначальное приспособление к усовершенствованию быстроты, вернее, длительности и большей легкости передвижения, очень знаменательно. Все древние растительноядные формы приспособлены к обитанию в областях болот и рек с концентрированным кормом. Поэтому быстрота и длительность передвижения не являются у них решающими для питания. Гораздо важнее сила в рытье при добыче подземных корневищ, преодолении топких мест или густой растительности. В этих древних биотопах долго удерживаются грубые, примитивные структуры скелета с тяжелыми, толстыми костями и малоподвижными суставами.
Размеры подобных зон с концентрированными кормами не могли быть очень велики, и приспособительная эволюция необходимо должна была обратиться к освоению более удаленных от воды сухих зон, где растительная пища была более бедной, особенно для древнего пресмыкающегося, не могущего доставать ее с деревьев. Кормовая концентрация на единицу площади здесь была гораздо ниже, и единственной возможностью для преодоления трудностей питания в новой области было энергичное передвижение. Обходы больших площадей при повышенной скорости обеспечивали животное пищей, но лишь при том непременном условии, что передвижение происходило при небольшой затрате энергии. Подобное движение невозможно для тяжелых архаических пресмыкающихся перми, нуждавшихся в богатой, скученно произрастающей кормовой растительности и обитавших только в узких ареалах, занимавших лишь небольшой процент пригодной для жизни площади материка.
Дицинодонты — самые многочисленные пресмыкающиеся верхней перми, с мощными челюстями и хорошо развитыми, очень сходными с примитивными млекопитающими, поясами конечностей, вначале отличались сильным развитием передних конечностей. В дальнейшем приспособлении к новым, широким кормовым пространствам у триасовых Rhadiodromus наблюдается быстрое развитие задних конечностей. Полное освоение открытых пространств с рассеянной пищей оказалось настолько сложным, что древние пресмыкающиеся — тероморфы — так и не смогли этого сделать. Слишком много структурных противоречий было в их архаических организмах. Новая группа — двуногие архозавры — повторили эту попытку, но окончательное завоевание сухих материковых пространств было достигнуто только поздними, неогеновыми группами млекопитающих и, главным образом, жвачными. В древних палеогеновых фаунах большинство млекопитающих повторяет, на другой биологической основе, приспособления к питанию в концентрированной кормовой зоне как требующие менее сложных приспособительных устройств.
Палеонтологический институт Академии наук СССР
Поступило 19 I 1951
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
1 И. А. Ефремов, ДАН, 20, № 2-3 (1938). 2 И. А. Ефремов, Тр. ПИН, 10, в. 2 (1940).
Источник:
Доклады Академии Наук СССР. 1951. Т. 77. № 3. С. 483-485.
